

Regulation of plant developmental transitions—age pathway
Plants undergo a series of developmental transitions during their life cycle. After seed germination, plants pass through two distinct phases: vegetative phase in which leaves are produced and reproductive phase in which flowering is occurred. Vegetative phase can be further divided into juvenile and adult phases, based on the reproductive competence and morphological changes. Growing lines of evidence indicate that microRNA156 (miR156) and its targets SQUAMOSA PROMOTER BINDING-LIKE (SPL) transcription factors, exert an evolutionarily conserved role in regulating developmental transitions in plants. Our lab is performing a comprehensive characterization of the age pathway, focusing on the regulatory basis of the developmental decline in miR156 level by age and the molecular mechanism by which miR156 regulates developmental transitions.

Key publications
[1] Yu S, Cao L, Zhou CM, Zhang TQ, Lian H, Sun Y, Wu JQ, Huang JR, Wang GD, and Wang JW*. (2013). Sugar is an Endogenous Cue for Juvenile-to-Adult Phase Transition in Plants. eLife. 2: e00269.
[2] Zhou CM and Wang JW*. (2013). Regulation of Flowering Time by MicroRNAs. Journal of Genetics and Genomics. 40: 211-215.
[3] Yu S, Galvao V, Zhang YC, Horrer D, Zhang TQ, Hao YH, Feng YQ, Wang S, Schmid M, and Wang JW*. (2012). Gibberellin Regulates Arabidopsis Floral Transition through miR156-Targeted SQUAMOSA PROMOTER BINDING-LIKE Transcription Factors. Plant Cell. 24: 3320-3332.
miR156-SPL as an Universal Timer in Arabidopsis
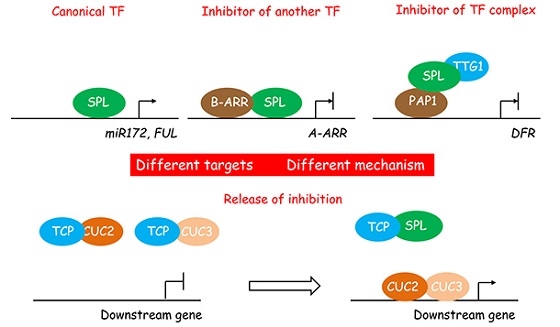
As in animals, age regulates diverse aspects of plant physiology and development. However, the underlying molecular mechanism is poorly understood. Overexpression of miR156 or target mimicry of miR156, which suppresses miR156 activity, results in pleiotropic phenotypes in plants, including A. thaliana, rice, maize, poplar and tomato. We are interested in how miR156 and its target SPL transcription factors regulate these diverse developmental and physiological processes.
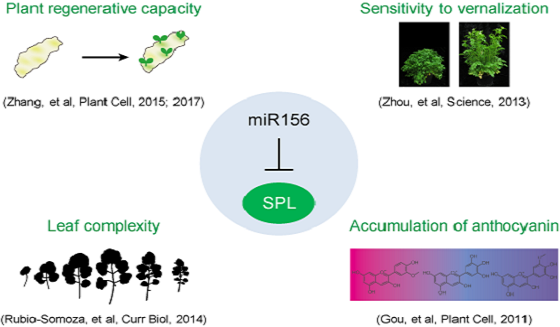
Key publications
[1] Zhang TQ, Lian H, Zhou CM, Xu L, Jiao Y, and Wang JW*. (2017). A two-step model for de novo activation of WUSCHEL during plant shoot regeneration. Plant Cell. DOI: 10.1105/tpc.16.0086.
[2] Zhang TQ, Lian H, Tang H, Dolezal K, Zhou CM, Yu S, Chen JH, Chen Q, Liu H, Ljung K, and Wang JW*. (2015). An intrinsic microRNA timer regulates progressive decline in shoot regenerative capacity in plants. Plant Cell. 27: 349-360.
[3] Rubio-Somoza I, Zhou CM, Confraria A, Martinho C, Born PV, Baena-Gonzalez E, Wang JW*, and Weigel D*. (2014). Temporal Control of Leaf Complexity by miRNA-Regulated Licensing of Protein Complexes. Curr Biol. 24: 2714-2719.
[4] Zhou CM, Zhang TQ, Wang X, Yu S, Lian H, Tang H, Feng ZY, Zozomova-Lihova J, and Wang JW*. (2013). Molecular Basis of Age-dependent Vernalization in Cardamine flexuosa. Science. 340: 1097-1100.
[5] Gou JY, Felipes FF, Liu CJ, Weigel D, and Wang JW*. (2011). Negative regulation of anthocyanin biosynthesis in Arabidopsis by a miR156-targeted SPL transcription factor. Plant Cell. 23: 1512-1522.
The molecular basis of perennial growth habit
The life cycle strategy of higher plants can be classified into three forms: annual, biennial and perennial. Under natural conditions, annual plants go from germination to death within one year. By contrast, biennial and perennial plants take at least two years to complete their biological lifecycles. Monocarpic perennials, such as bamboo, bloom only once and subsequently die, whereas polycarpic perennials continue to flower in subsequent years (although not necessarily every year). To dissect the molecular mechanism of seasonal flowering in perennial, we used bitter cress (Cardamine flexuosa), a herbaceous Brassicaceae widely distributed in Europe, and Populus x Canadensis (poplar).
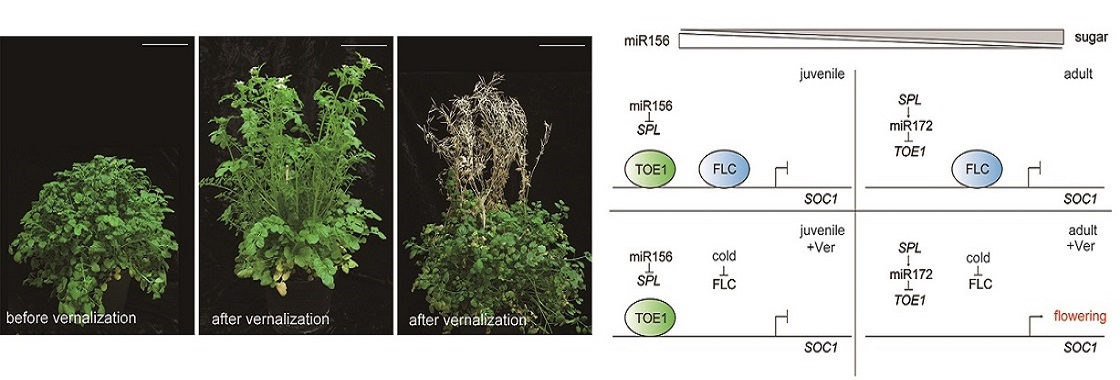
Recently, we demonstrate that the level of miR156 sets a threshold for vernalization response. Age and vernalization pathways coordinately regulate flowering through modulating the expression of CfSOC1, a flower-promoting MADS-box gene. We note that, although both vernalization and age pathways operate in the related annual Arabidopsis thaliana, this species does not confer an age-dependent vernalization response. Thus, our results suggest that the recruitment of age cue in response to environmental signals contributes to the evolution of life cycle in plants.
Key publications
[1] Zhou CM, Zhang TQ, Wang X, Yu S, Lian H, Tang H, Feng ZY, Zozomova-Lihova J, and Wang JW*. (2013). Molecular Basis of Age-dependent Vernalization in Cardamine flexuosa. Science. 340: 1097-1100.
[2] Wang JW*, Park MY, Wang LJ, Chen XY, Weigel D, and Poethig S*. (2011). MiRNA Control of Vegetative Phase Change in Trees. PLoS Genet. 7: e1002012.